浅说动力学模型(下篇)
原创: 赵洪龙 宇宙实验媛
图1. 数学建模在生物学研究中的应用范式
书接上文浅说动力学模型(上篇),由于本人主要研究植物学代谢建模,下面就举例说明使用模型在植物生理研究中取得的一些重要成就。在过去的研究中,植物领域对光合作用的研究积累了大量的数据,这为光合作用相关的动力学建模提供了数据基础。研究者们已经从细胞代谢层面(1)、叶片层面(2, 3)、植株水平和群体水平分别构建了模型(4)。 利用这些模型已经成功预测出了限制光合作用的关键过程,并且部分预测结果已经通过基因工程改造进行了验证。
自然条件下,植物生长的环境总是不断变化的,比如一阵微风吹过就会导致植物叶片接收的光发生改变。上层叶片摆动,下层叶片就会处于一明一暗的不断变化中。正如人眼睛的瞳孔在黑暗条件下放大、在明亮条件下会缩小,而闪光对眼睛有很强的刺激作用一样,闪光对植物而言也是一种非常严重的胁迫,会导致光合作用电子传递链出现损伤,从而抑制光合作用。植物有很多机制能耗散过剩的能量和修复这种闪光带来的电子传递链损伤,以防止被‘烧伤’并最大化光能的利用效率。以前没有一个合适的系统模型评估过这种能量耗散途径对作物产量所造成的损失,故而也一直没有明确的方案能通过减少这种损失来提高作物产量的潜力。Zhu等构建的作物群体中叶片光合作用能量分配的动力学模型,结合光线追踪算法对这个过程进行了理论分析。其预测显示,如果能够加快这个修复过程,作物的生产能力能增加20%左右(5)。
为了能更好地理解这个原理,我们给出了示意图(图2)。叶片在强光条件下的时候,一般来说植物的光合能力相对要高一些,而过多的能量会通过其他的途径耗散,以免叶片吸收过多的能量无法被转化成化学能导致叶片‘烧伤’。这对大部分植物叶片来讲是一种很好的防御机制(叫做光损伤防御)。然而,当植物从强光条件下转移到低光条件下的时候(t0时刻),能量往往成为光合能力的一种限制条件。此时,那些防御途径仍然“大手大脚”地耗散光能,减少了分配到化学能转化的分量。在数分钟或者数小时(假设为t0 – t2)的低光下,这些防御途径耗散的能量会慢慢减少,光合同化能力慢慢增加,我们把这段时间称之为恢复时间。就像我们从太阳底下走进相对比较暗的房间一样,眼睛会突然感觉到很暗,但过一段时间后房间里面就会变得‘亮起来’。从图2中我们可以很容易理解,如果减少这段恢复时间,就能够增加总的光能利用效率。因此,Zhu等2004年建立的模型主要描述了:
**1. 植物冠层中上下不同层次的点在一天的光强变化条件下的太阳辐射截获量;
- 截获的太阳辐射与光合能力之间的对应关系;
- 接受的太阳辐射量变化时,植物由于光损伤防御机制的存在而引起的能量损失(及温度的影响);
- 通过减少光损伤防御恢复时间能给作物的光合生产能力提升带来的潜在空间。
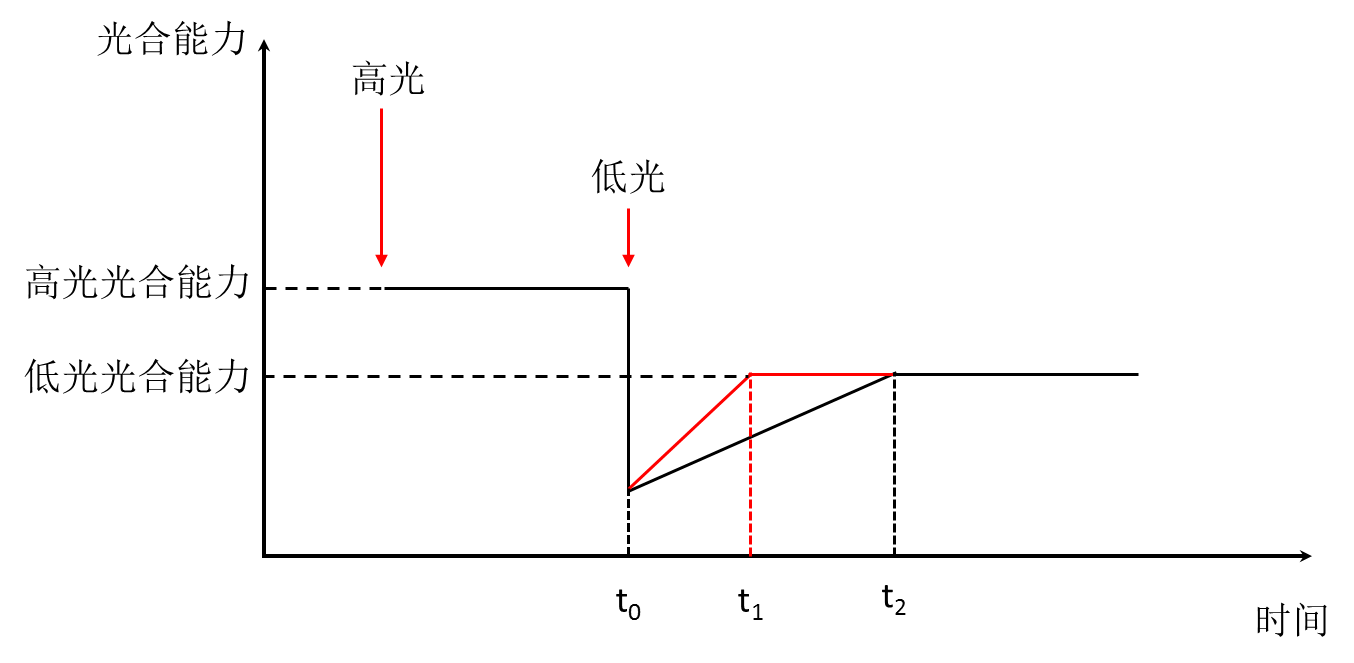
图2. 植物叶片从强光变到至弱光条件下时光损伤防御恢复机制的简单示意图。红色箭头表示叶片暴露在不同的光下。如果能把光损伤防御恢复的时间从t2缩短至t1,将有可能提高整个群体光合生产能力的12-30%。历经12年其合作团队通过基因工程手段进行改造,加快了这种光损伤恢复的速度,实现了烟草的光合作用能力及生产能力的大幅增加,成功验证了这一预测(6)。不仅如此,Zhu等建立的卡尔文循环代谢模型结合进化算法,预测了光合作用碳同化代谢过程中限制其效率的关键酶,这些预测与报道的实验结果一致(1),因此也被采纳为作物改良的主要策略之一(7, 8)。Wang等建立的C4光合作用代谢模型系统解析了C4光合作用代谢过程高效的互作机理(9)。 利用动力学模型,还可以为将来合成生物学导入新的代谢途径提供理论分析工具。** 比如通过参考模型计算和分析的结果进行基因工程改造,增加微生物中特定代谢物的产量,这在微生物研究的应用中较为广泛(10);或者增加植物的次生代谢产物的产量(11) 等。
有关复杂代谢系统生物学建模的方法,Zhu(12)和Zhao(13)等均分别进行了综述,感兴趣的读者可参考阅读。
展望动力学模型虽然近些年来,随着数据测量技术和计算能力的飞速提升,系统生物学的发展势头迅猛,也取得了一些成就,然而现在动力学模型的构建过程中依然存在一些挑战。首先,动力学模型是基于具体过程的一类模型。而生物学过程总是交织成多维度的复杂网络,针对具体问题解析这些关系是一件繁琐的事情,因此模型构建是一件十分耗时的工作,且对研究的问题需要有深厚的知识背景,况且,现在我们并没有完全了解所有的生命过程。更甚,对于一个模型的参数化和验证而言,难以收集到一套完整而系统的数据集。数据库和文献中收集的数据均非常离散,环境、物种和测量过程的差异都会导致模型参数化和验证过程陷入困境。第三,模型参数化的过程和方法需要进一步提升并规范化。
笔者认为,决定未来动力学模型构建与生物学研究耦合的几个重要因素如下:
- 第一,生物学测量技术需要进一步发展,高通量、高标准和高质量的数据是模型构建的基石。虽然目前组学数据的增长速度史无前例,但测量方法之间的差异、规范性问题、条件和细胞特异性问题等依旧是建模的重要拦路虎。
- 第二,多背景交叉融合需要进一步加强。生物学与化学、物理学、数学、计算机科学等学科之间的交流能在确保重要信息的前提下更好地将复杂生物学问题简单化,从而确保模型具有更强的生物学预测能力和应用价值。
- 第三,模型需要不断被优化和更新。所有的模型都不可能是完美的,需要针对具体问题进行结构和参数的进一步优化,因此需要构建一些大型的数据存储和模型分析平台,从而进入用模型帮助数据分析、用数据优化和修正模型的良性循环。至此,方能更好地将模型应用于生物学研究,造福于人类。
参考文献:
Zhu XG, de Sturler E, & Long SP (2007) Optimizing the distribution of resources between enzymes of carbon metabolism can dramatically increase photosynthetic rate: a numerical simulation using an evolutionary algorithm. Plant physiology 145(2):513-526.Tholen D & Zhu XG (2011) The mechanistic basis of internal conductance: a theoretical analysis of mesophyll cell photosynthesis and CO2 diffusion. Plant physiology 156(1):90-105.Xiao Y, Tholen D, & Zhu XG (2016) The influence of leaf anatomy on the internal light environment and photosynthetic electron transport rate: exploration with a new leaf ray tracing model. Journal of experimental botany 67(21):6021-6035.Song Q, Chu C, Parry MA, & Zhu XG (2016) Genetics-based dynamic systems model of canopy photosynthesis: the key to improve light and resource use efficiencies for crops. Food Energy Secur 5(1):18-25.Zhu XG, Ort DR, Whitmarsh J, & Long SP (2004) The slow reversibility of photosystem II thermal energy dissipation on transfer from high to low light may cause large losses in carbon gain by crop canopies: a theoretical analysis. Journal of experimental botany 55(400):1167-1175.Kromdijk J, et al. (2016) Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 354(6314):857-861.Miyagawa Y, Tamoi M, & Shigeoka S (2001) Overexpression of a cyanobacterial fructose-1,6-/sedoheptulose-1,7-bisphosphatase in tobacco enhances photosynthesis and growth. Nat Biotech 19(10):965-969.Simkin AJ, McAusland L, Headland LR, Lawson T, & Raines CA (2015) Multigene manipulation of photosynthetic carbon assimilation increases CO(2) fixation and biomass yield in tobacco. J Exp Bot 66(13):4075-4090.Wang Y, Brautigam A, Weber AP, & Zhu XG (2014) Three distinct biochemical subtypes of C4 photosynthesis? A modelling analysis. Journal of experimental botany 65(13):3567-3578.Colón AM, Sengupta N, Rhodes D, Dudareva N, & Morgan J (2010) A kinetic model describes metabolic response to perturbations and distribution of flux control in the benzenoid network of Petunia hybrida. The Plant Journal 62(1):64-76.Xin CP, Tholen D, Devloo V, & Zhu XG (2015) The benefits of photorespiratory bypasses: how can they work? Plant physiology 167(2):574-585.Zhu XG (2010) Systems-level modeling–a new approach for engineering efficient photosynthetic machinery. Journal of biotechnology 149(3):201-208.Zhao H, Xiao Y, & Zhu XG (2017) Kinetic Modeling of Photorespiration. Methods Mol Biol 1653:203-216.



